 |
|
ファザール・ラナ Facts for Faith quarter 4, 2001, Issue 7, 33-41 松崎英高(箱崎キリスト福音教会牧師)、ティモシー・ボイル(つくばクリスチャンセンター宣教師、物理学学士号、神学博士)共訳 人間は、数百万年以上も這い回る段階を経て存在するようになったのでしょうか。それとも、人間は、超自然的なデザインによって二足歩行へと飛躍し たのでしょうか。人類は、アメーバから生じたのでしょうか。それとも、創造者が生命に目的や価値や意味を持たせようと意図されたのでしょうか。そのよう な質問に対し如何に答えるかは、人間社会が最も切迫している問題への対処の仕方に大きく影響します。人間が神によってデザインされた、繊細な感覚を持 つ、霊的な被造物なら、自然の気まぐれな偶然により生じた場合よりも大きな配慮と考慮を払う価値があります。 ある科学者たちは、人間とはまれにしか起こらない運命の気まぐれであり、過去500万年にわたって起こった偶然の出来事によって存在するように なった知性のある猿であると言います。彼らは、現生人類に至る過程において役割を果たしたのは自然選択であると主張します。生存競争と捕食動物と環境か らの圧力が遺伝的な変化を徐々に選択していったのだと、彼らは言います。これらの変化は生存率と繁殖の成功率を向上させたと推定されます。このようにし て、自然選択は度々偶然に出現した変異に作用して、連続的に新しい種を作りつつ、ついには偶然によって現生人類が出現したのだろうとされます。 大きなサイズの脳(実際には、全体重に対する脳の比率)と手先の器用さと文化の発展と並んで、二足歩行は最も重要な人間の特徴の一つです。進化論 的な立場に立つ人類学者にとって、二足歩行の出現と発達を理解することは、人間の起源を理解することと同等であるとされます。 進化論的な思想とは著しい対照をなすものですが、聖書は、人間を神の創造行為の頂点にあるものとし、神のイメージに創造され、他のすべての被造物 とは区別されるものであることを啓示しています[1]。人類の起源についての聖書の説明には、人間を創造するために神がサルから人間への進化的な変化過程を利用したと考 える余地はまったくありません。初めから計り知れない価値を持ち、また肉体を持ちつつも霊的な存在でもある最初の人間たちを創造するに際して、神の直接 的な関与があったと、聖書は描いています。 人間の起源に関しての聖書と進化論による説明(の真偽)を判別するためには、それぞれを検証できるかどうかが重要なポイントになります。それを可 能にする強力で新しいアプローチがあるのです[2]。(脚注を参照のこと)起源に関するこれらのモデルによって予測されるものは、厳格な科学的な検証に供することがで きます。科学的な記録により最大の支持を受け、かつ新しい発見とも最も良く調和するものが、最も正確なシナリオであると実証されます。 二足歩行に関連する古生物学(二足歩行する霊長類の化石記録に関する研究)と古生態学(古代の環境に関する研究)における最近の発展は、データ比 較を実施する異例の機会を与えてくれます。他の専門分野でもそうですが、この分野における新しい発見は、二足歩行の自然主義的な起源説と相反するもので すが、聖書の記録の信憑性を十分に確認させてくれます。 進化論的なシナリオ進化論的なパラダイムに従えば、類人猿に似た祖先から類人猿と人類の両方の系統が生じたので、二足歩行する霊長類はナックル歩行(指背歩行)をす る四足動物から進化したに違いありません。ナックル歩行とは、チンパンジーやゴリラが有する陸上の四足歩行(四肢のすべてを使って地上を移動すること) の特殊なタイプです[3]。ナックル歩行をする動物は、手のひらや指にではなく、指関節に体重をのせます。このような構造は、チンパンジーと ゴリラが木に登ったり、ぶら下がったりするために長く湾曲した器用な指を持ちながらも、四肢のすべてを使って歩行することを可能にします。 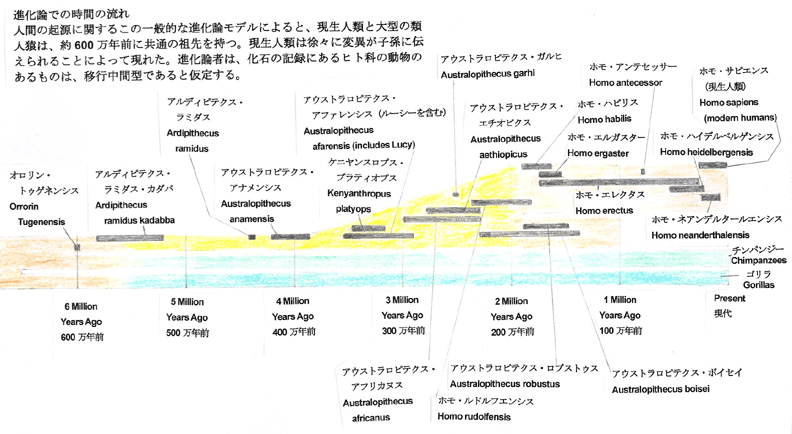 古人類学者たちは、自然的なプロセスによってどのようにして二足歩行が現われるようになったのかを説明するために、無数の仮説を提案します。初期 の説明の一例を挙げると、二足歩行は道具使用のため、手を自由に利用するために生じたというものがありました。化石の記録がこの意見に相反するもので あったので、進化論的な生物学者たちはこの考えを否定しました。考古学的な記録は、道具の使用が出現する少なくとも200万年前に二足歩行が存在したこ とを明白にしています[4]。 二足歩行の出現を説明するために模索しつつ提出された仮説のほとんどは、東アフリカの環境が森林地帯から乾燥した見通しが利くサバンナに変わった ことが前提になっています[5]。そのような変化に伴って、陸生の四足動物は食料が不足し、捕食動物の餌食になる危険が増大し、直射日光を避けるこ とができなくなりました[6]。 二足歩行は、このような問題に対処するための手段となります。直立歩行は、遅いスピードで移動するにはエネルギー効率がより優れた方法です[7]。このことは、二足歩行の霊長類が食料を捜し求めて長い距離を行き来することを可能にしました。ひとたび食料を見つ けたなら、彼らは自らの子供たちのために用意された“ねぐら”に戻る際に、遠距離からその食料を運ぶことができました[8]。 二足歩行する霊長類の頭の高さは、見通しの利くサバンナでは、四足動物の類人猿と比較して、捕食動物の餌食になることをより効果的に避けることが できます。直立姿勢は、二足歩行する霊長類が直ちにそしてより遠距離から、捕食動物を探知することを可能にしたことでしょう。 二足歩行はまた、体温調節をするに際にも有利です[9]。直立して二足歩行する霊長類の熱吸収は、四肢のすべてを使用して歩く類人猿より60%も少ないのです。四足動物の 姿勢は背中全体を直射日光にさらしますが、直立姿勢は頭部と肩をさらすだけです。 進化論的な生物学者たちには、霊長類に二足歩行をさせた淘汰の圧力に関する意見の一致がなく、また、容認された時間(次の項を参照のこと)でその ような劇的な変化をもたらしたメガニズムを証明するものもありません。今までのところ、四足歩行から二足歩行への変化の背後に働いた進化の圧力について の唯一の合理的な要因として、東アフリカ全体に及んだ森林地帯という生育環境の消失だけが残っています。 二足歩行の解剖学ナックル歩行する四足動物から直立の二足動物へ移行することは、大規模な解剖学的な変化が必要です[10]。これらの変化には次のものが含まれます。
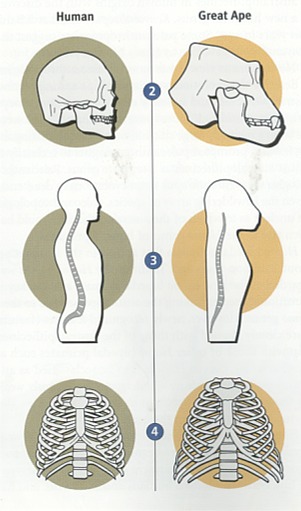 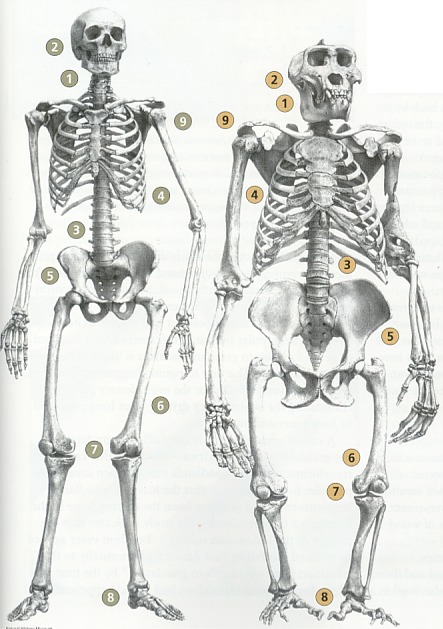 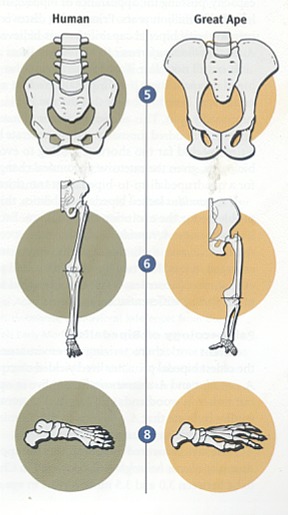 予測ナックル歩行する四足動物が二足歩行する霊長類に変化するために必要な劇的な解剖学的な変化が、どのようにして起こり得たのかを想像する試みでさ え、挫折させられます。にもかかわらず、二足歩行が自然的プロセスによる生物学的進化によって生じたと仮定するなら、類人猿と人類が分岐して後に、二足 歩行が段階的に生じて、次第にはっきりと現われるようになったはずです。さらに、最初に現われた二足歩行の形は、未完成で非効率なものであったはずで す。一旦現われたとなると、二足歩行は現生人類のより効率の良い二足歩行に段階的に移行して行ったはずです。その上、もし本当にそのような変化が起こり 得たのなら、環境とライフスタイルに完全に適合していたナックル歩行する霊長類が直立歩行する霊長類に変化するためには、十分な進化の圧力が必要となり ます。 最近の科学的な発展化石と地質学的な記録における最近の発見は、二足歩行の起源と自然史に関する古人類学者たちの見解を抜本的に変えてしまいました。これらの新しい 科学的な発展は、進化論的なシナリオを拠り所とする予測と全く矛盾しています。 二足歩行の最初の出現1994年と1995年に、古人類学者たちは二組の発見を報告しました。それらの報告には、アウストラロピテクスに属する二つの種の化石について 記載されていました。一つの研究チームは、エチオピアで440万年と年代測定されたヒト科の動物の化石を発見しました[11]。この種を、彼らはアウストラロピテクス・ラミダスと命名しました。後にそれはアルディピテクスという新しい属 に再分類されました[12]。 その一方で、別の研究チームは、ケニアでヒト科の化石を一体発見し、390万年前から420万年前と年代決定しました[13]。これらの種は、アウストラロピテクス属と認知され、アウストラロピテクス・アナメンシスと命名されました。さ らに調査が続けられて、その年代は407万年前と確認されました[14]。A.アナメンシスの脛骨の分析はその二足歩行の能力をはっきりと証明し、二足歩行の出現を少なくともさらに 50万年過去に押しやりました。この発見の前には、二足歩行の能力を持った最古の霊長類は、アウストラロピテクス・アファレンシス(約390万年前)で あると信じられていました。 アウディピテクス・ラミダスが二足歩行の能力を持っていたかどうかは、まだ定かではありません。もしそうだとすると、二足歩行の最初の出現は、類 人猿と人類の系統が分かれたとされる時期に大変接近しています。このことは、自然選択の力はたったの数十万年で二足歩行を出現させたことになります。四 足歩行から二足歩行への移行に必要な大規模な解剖学的な変化を考えると、進化論的な生物学者たちの主張に沿えば、この期間はあまりにも短すぎます。 もしA.ラミダスに二足歩行の能力が欠如していたのなら、このこともまた進化論的なパラダイムにとっては障害となります。進化論的な生物学者たち は、A.ラミダスをA.アナメンシスに至る祖先種とみなしています。このシナリオによれば、二足歩行は20万年以下で生じたことになります。それは、莫 大な種分化が起こるにはさらに短い時間です。従って、そのようなことが実現した可能性はさらに少なくなります。 二足歩行の古生態学最古の二足歩行の霊長類が生活した環境を特定する最近の研究によって、予期せぬ結果が得られました。A.ラミダスとA.アナメンシスは見通しの利 くサバンナにではなく、森林地帯に住んでいました[15]。さらに、最近の研究では、A.アファレンシスは森林地帯と見通しの利くサバンナが入り混じった環境に住んでい たことが指摘されています[16]。 新たに発見されたアウストラロピテクス属であるアウストラロピテクス・バーレルガザーリはチャドで収容され、300万年から350万年前と年代決 定されました。それもまた、森林地帯とサバンナの入り混じった居住環境に住んでいました[17]。そして、新たに発見されて、350万年前と年代決定されたヒト科の動物であるケニアントロプス・プラティオプ スは、見通しが利く草原を含むが、森林地帯が優勢であるような環境に住んでいました[18]。 人類学者であり、サイエンスライターでもあるロジャー・ルーウィンの言葉を引用すると、「密林から草原のサバンナに大股で歩いて出て行く我々の祖 先についての有名な概念は事実ではなく、作り話であるようです[19]。」ルーウィンによって表現されたこの新しい認識は、進化論的なパラダイムにとっては深刻な障害となります。と いうのは、二足歩行を出現させたと長い間にわたって予想されていた進化の推進力が排除されたからです。 森林地帯から見通しが利くサバンナに移行する原因となった東アフリカの乾燥化を理解するためになされた最近の地質学的な研究は、森林地帯という環 境の喪失が二足歩行を出現させる推進力ではなかったという証拠をさらに提供します。この研究は、インドネシア海路の閉鎖が東アフリカの降雨量を減少さ せ、その結果、森林地帯が草原地帯に変化したのは300万年から400万年前であったことを指摘しています[20]。東アフリカが乾燥するまでには、二足歩行はすでに出現していました。 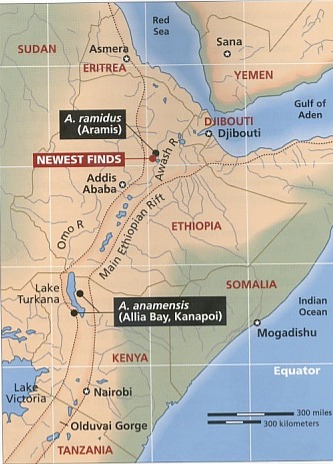 変化しない二足歩行類人猿と明白なヒト科の動物と現生人類の200を超える骨盤の標本について最近行われた数学的及び統計的分析は、二足歩行の出現を進化論的に説明 することの正当性をその根幹において疑わせる、歴史的なパターンを明らかにしています[21]。二足歩行は長い時間を掛けて徐々に変化するのではなく、突然現われると長い期間静止したまま(変化のないま ま)であり、それから、突然の変化が起こります。そして、再び静止した状態になり、その後に急速な変化が起こるのです。 最初の二足歩行する霊長類であるアウストラロピテクスは、ヒト属の霊長類とは区別される二足歩行の型を持っていました。アウストラロピテクスは、 条件的な(選択が自由な)二足歩行をしましたが、ヒト属は絶対的な二足歩行を常に行っていました。アウストラロピテクスは殆ど300万年間存在しました が、彼らの二足歩行はヒト属の絶対的な二足歩行に徐々に変化しませんでした。むしろ、アウストラロピテクスが存在する期間、それはずっと静止したままでした。 ヒト属の出現とともに、絶対的な二足歩行が突然、化石記録に現われます。進化論的な観点からは、この急速な変化には条件的な二足歩行から絶対的な 二足歩行に至る急速な移行過程が必要です。ヒト属の絶対的な二足歩行は約200万年間静止したままです。興味深いことに、ホモ・エレクトスとネアンデル タール人は同一の二足歩行の型を持っていましたが、それは現生人類に見られるものとははっきりと区別される型でした。現生人類の出現とともに、二足歩行 の別の型が突然に現われ、その時から今日までそれが継続しています。化石記録に見られる突然の変化によって中断される静止状態というパターンは、進化論 的な予測を裏切るものですが、神の創造行為の明らかな指標となります。もし神がアウストラロピテクスや二足歩行するヒト属や類似した他の属を創造された のなら、それぞれの属が所有する二足歩行は、それが生きた環境とライフスタイルの状況に最適化されているはずだと予測できます。このような創造が一旦な されると、どのような変化も二足歩行の最適化された型から外れて最適性を危うくするので、自然選択は二足歩行のそれぞれの型を静止したままにすると予想 されます[22]。さらに、ライフスタイルと環境の違いを考えると、化石の記録に見られるように、二足歩行の異なる型を持つアウ ストラロピテクスとヒト属を神が創造されたことは、創造モデルの観点から導かれるのです。 結論二足歩行の自然史における最近の科学的な発展により、有用な観察結果が収集されたことが、人類の起源に関しての進化論的なシナリオと聖書的なシナ リオの両方を評価することを可能にしました。化石記録における初期の二足歩行が突然に出現しているということは、二足歩行が自然のプロセスである生物学 的進化によって出現するための十分な時間がなかったことを意味します。また、化石記録は、原始的な二足歩行からさらに複雑で効果的な型へと段階的に移行 したというパターンを描けないでいます。これらの変化を押し進めるどのような有意な進化の圧力も存在しなかったことが、二足歩行の出現をさらに目を見張 るものにします。 二足歩行の型の急激な変化によって分断された二つの静止期を伴って、初期の二足歩行が突然に出現していることは、二足歩行の自然史の特徴です。こ れらの特徴は、特殊創造が予想するパターンと完全に合致しています。 神が二足歩行する大型の霊長類を創造したとする聖書的な創造モデルは、長い期間の静止期を予測します。すなわち、完全無欠の創造者は、その動物が さらされる環境や捕食動物の脅威や激しい生存競争という難問に理想的に適合する二足歩行の型を用意されたと思われます。最近の科学的発見は、人間の最も 重要で決定的な特徴の一つである二足歩行が神の直接的な創造の業に由来することを支持する明快な証拠を与えてくれます。人間以外の二足歩行する霊長類 は、特定の目的と機能を果たすためにデザインされました。彼らは創造者の作品でした。 二足歩行する霊長類に対してこのような神の配慮があったことを支持する証拠を伴って、神のイメージに独自に創造された人間の威光は、途方もなく大 きな意義を持ちます。人間が配慮に富む神によって目的をもって創造されたということは、人生を価値あるものにします。そのような意味合いを理解する社会 は、そこに暮らす人々に価値と意味と目的を与えます。また、そのような理解は、二足歩行への飛躍に関連する発見を極めて貴重なものであると認識させま す。 ラミダス猿人に関する最新情報この記事を出版するための最終段階に入ってから、カルフォルニア大学バークレー校の考古学者のチームが520万年前から580万年前と年代測定さ れたヒト科の化石の発見を報告し、この動物の環境を記述しました[23]。彼らの研究の成果は、二足歩行性が超自然的に出現したという主張を支持します[24]。 この発見をした古人類学者たちは、この化石をアルディピテクス・ラミダスと分類しました。その分析の結果は、A.ラミダスが直立歩行をしていたこ とを明らかに示すものでした。この劇的な発見は、ヒト科の化石記録をほとんど100万年遡らせるだけでなく、二足歩行性の出現がヒト科の動物が最初に出 現した時期と一致することを意味するのです。二足歩行性は化石記録の中にまさしく突然に出現するのです。 また、古人類学者たちは、A.ラミダスがもっぱら湿潤した森林という環境に住んでいたことを割り出しました。同様に、440万年前と年代測定され たA.ラミダスも湿潤した森林を生育環境にしていました[25]。これらの発見は、二足歩行性を出現させるための進化の推進力を完全に排除するものです。ある研究者が論評した ことですが、これらの発見は、「ヒト科の進化の方式と時期について長年抱いてきた考えに意義を申し立てているのです[26]。」 引用文献1 Genesis 1:26-27; Genesis 2:7; Genesis 2:22; Mark 10:6; Matthew 19:4; Psalm 8:4-5. 2 Hugh Ross, “Can Science Test a ‘God-Created-It’ Model? Yes!” Facts for Faith(Q2 2000), 40-47; 55-58. 3 John G. Fleagle, “Primate Locomotion and Posture,” in The Cambridge Encyclopedia of Human Evolution, paperback edition, ed. Steve Jones, Robert Martin, and David Pilbeam (New York: Cambridge University Press, 1994), 75-85. 4 Eric Delson et al., eds., Encyclopedia of Human Evolution and Prehistory, 2d ed. (New York: Garland Publishing, 2000), 394-95; B. Bower, “African Fossils Flesh Out Humanity’s Past,”Science News155 (1999), 262; Elizabeth Culotta, “A New Human Ancestor?”Science284 (1999), 572-73; Jean de Heinzelin et al., “Environment and Behavior of 2.5 Million-Year-Old Bouri Hominids,”Science284 (1999), 625-29; Berhane Asfaw et al., “Australopithecus garhi: A New Species of Early Hominid from Ethiopia,”Science284 (1999), 629-35. 5 Roger Lewin, Principles of Human Evolution: A Core Textbook(Malden, MA: Blackwell Science, 1998), 219-22. 10 Lewin, 218; Robert Martin, “Walking on Two Legs,” in The Cambridge Encyclopedia of Human Evolution, paperback edition, ed. Steve Jones, Robert Martin, and David Pilbeam (New York: Cambridge University Press, 1994), 78; Fred Spoor et al., “Implications of Early Hominid Labyrinithine Morphology for Evolution of Human Bipedal Locomotion,”Nature369 (1994), 645-49. 11 Tim D. White et al., “Australopithecus ramidus, a New Species of Early Hominid from Aramis, Ethiopia,”Nature371 (1994), 306-12; Henry Gee, “New Hominid Remains Found in Ethiopia,”Nature373 (1995), 272. 12 Tim D. White et al., “Corrigendum,” Nature375 (1995), 88. 13 Meave G. Leakey et al., “New Four-Million-Year-Old Hominid Species from Kanapoi and Allie Bay, Kenya,” Nature376 (1995), 565-71. 14 Meave G. Leakey et al., “New Specimens and Confirmation of an Early Age for Australopithecus anamensis,”Nature393 (1998), 62-66; B. Bower, “Early Hominid Rises Again,”Science News153 (1998), 315. 15 Meave Leakey and Alan Walker, “Early Hominid Fossils from Africa,” Scientific American(June 1997), 74-79; Clark Spencer Larsen, Robert M. Matter and Daniel L. Gebo,Human Origins: The Fossil Record, 3d ed. (Prospect Heights, IL: Waveland Press, 1998), 46. 17 Michel Brunet et al., “The First Australopithecine 2,500 Kilometers West of the Rift Valley (Chad),” Nature378 (1995), 273-75. 18 Meave G. Leakey et al., “New Hominid Genus from Eastern Africa Shows Diverse Middle Pliocene Lineages,” Nature410 (2001), 433-40. 20 Mark A. Cane and Peter Molnar, “Closing of the Indonesian Seaway as a Precursor to East Africa Aridification Around 3-4 Million Years Ago,” Nature411 (2001), 157-62. 21 Francois Marchal, “A New Morphometric Analysis of the Hominid Pelvis Bone,” Journal of Human Evolution38 (2000): 347-65. 22 Niles Eldredge, Reinventing Darwin: The Great Debate at the High Table of Evolutionary Theory(New York: John Wiley, 1995), 78-81. 23 Yohannes Haile-Selassie, “Late Miocene Hominids from the Middle Awash, Ethiopia,” Nature412 (2001), 178-81; Giday WoldeGabriel et al., “Geology and Paleontology of the Late Miocene Middle Awash Valley, Afar Rift, Ethiopia,”Nature412 (2001), 175-78. 24 Henry Gee, “Return to the Planet of the Apes,” Nature412 (2001), 131-32; Michael Balter and Ann Gibbons, “Human Evolution: Another Emissary from the Dawn of Humanity,”Science293 (2001), 187-89. 25 Giday WoldeGabriel et al., “Ecological and Temporal Placement of Early Pliocene Hominids at Aramis, Ethiopia,” Nature371 (1994), 330-33. 26 Balter and Gibbons, 187-88. |